Student JC: Chronic circadian disruption modulates breast cancer stemness and immune microenvironment to drive metastasis in mice
Welcome to our Journal Club! I use this as a forum for open discussion about the paper in question. Anyone can participate in the journal club, and provide comments/critiques on the paper by leaving a comment below. This is the first post written totally by a guest author, lab member Leah Boyd! I wanted to re-start this journal club with posts from students in the lab, so hopefully this will be the first of many posts to come.
We all know that the effects of jetlag are pretty miserable—the fatigue, bizarre sleep schedule and desire for meals at odd hours of the day come with every transmeridian flight. But in 2019, Eva Hadadi, Hervé Acloque and colleagues at various French universities found that chronic jetlag and circadian rhythm disruptions (CRD) can moderately affect primary tumor development and significantly increase cancer-cell dissemination and metastasis in breast cancer. Their paper, entitled Chronic circadian disruption modulates breast cancer stemness and immune microenvironment to drive metastasis in mice, gives a good overview of what they found.

(Credit: Bob Al-Greene/Mashable)
Previous research has alluded to correlations between circadian rhythms and cancer development, and the International Agency for Research on Cancer classified CRD as a probable carcinogen in 2007. Van Dycke et al. performed a study in 2015 that showed p53 mutant mice (with p53 only deleted in the mammary gland) who experienced CRD developed mammary tumors eight weeks earlier than they typically would. But what happens after the tumors develop?
Hadadi et al. wanted to study beyond tumorigenesis and see how CRD affects tumor progression, cancer-cell dissemination and immune phenotype. They used the MMTV:PyMT model of spontaneous murine mammary carcinogenesis to test the impact of chronic CRD at the beginning of puberty-initiated tumorigenesis.
To start off, Hadadi et al. divided mice carrying MMTV:PyMT and MMTV:LUC transgenes (which express bioluminescent luciferase to tag the PyMT+ tumor cells) into two groups when they were at 6 weeks. For 10 weeks, the first group, which I’ll call the control group, experienced normal light and dark periods—12 hours of light, 12 hours of dark—while the other group faced an 8 hour reduction in the dark period every other day, which was designed to mimic the effects of night shift work or recurrent eastbound transmeridian flights. (Why does this matter? Previous research suggests that circadian rhythm is more altered by advances rather than delays in time.) Using in vivo imaging, they found that there was no major difference in the onset of tumorigenesis between the two groups but that the tumor burden and growth was significantly higher in the jetlag mice. Additionally, the lesions in the jetlag mice were more malignant (moved around the body more), even though the researchers observed multiple tumor grades in the primary tumors of both groups (Figure 1).
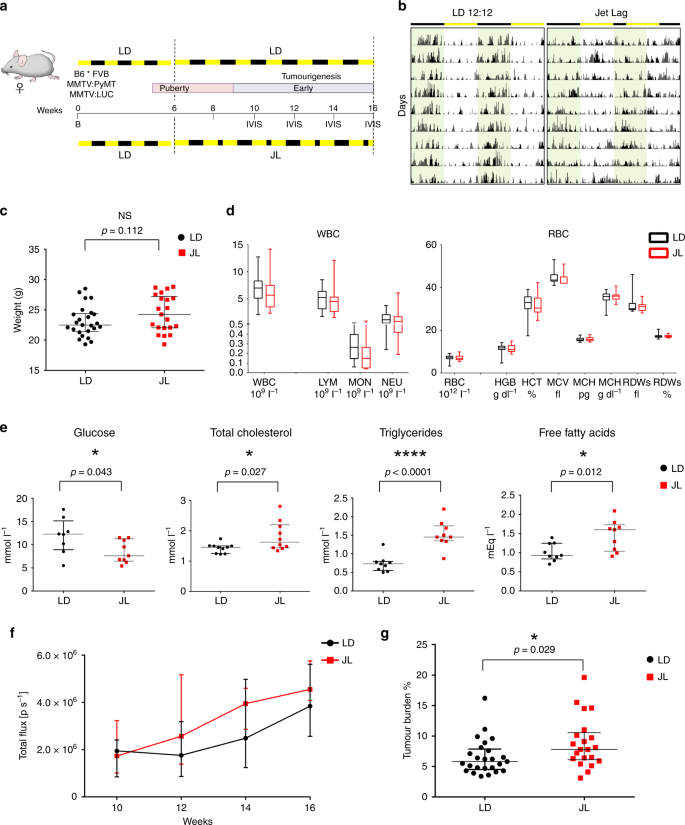
Figure 1: a. Experimental timeline for evaluation of the effect of chronic jet lag on spontaneous mammary tumorigenesis in B6*FVB PyMT mice. b. Tumor growth monitoring using bioluminescence. c. Weight at sacrifice of mice in LD and JL conditions. d. Blood cell counts: total numbers of white blood cells and red blood cells in LD and JL mice. f. Timeline of tumor growth in total flux measured by in vivo bioluminescence imaging in LD or JL groups. g. Tumor burden (tumor to body weight ratio) as % in LD or JL conditions. (Source: Hadadi et al., 2019)
The researchers used the same mice to explore if chronic CRD affected cancer-cell dissemination. They found a significant elevation of transgene expression in the bone marrow of the jetlag mice; in fact, the jetlag mice had an almost two-fold increase in disseminating cancer cells (DCCs) in the bone marrow. Flow cytometry analysis analysis showed an increase of circulating cancer cells in the bloodstream of the jetlag mice. Bone lesions demonstrated that cancer cells were further disseminating to bone (Fig. 2). Metastasis was much more prevalent in the jetlag mice (52%) than in the control group (28%), a result consistent with the aforementioned findings. CRD clearly promotes metastasis-but what elevates the risk for metastasis? (Clock genes. You’ll find out more later.)
A hierarchical clustering analysis on five jetlag and five control mice showed that a few genes were significantly differentially expressed between the two groups. But the genes with the greatest differential expression, including Rhodopsin and Gnat1, weren’t related to circadian cycles at all; they were actually linked to photoperception and phototransduction. Interestingly, they were downregulated in the primary tumor and mononuclear bone marrow cells of jetlag mice compared to the control group. The researchers noted that it’s unclear whether this finding was because of (a) the circadian rhythms of the control mice or (b) the peripheral tissues play a genuine functional role in phototransduction (or another unknown process these genes control). It will require deeper investigation by researchers.

Figure 2. a. Representative gating strategies for mammary stem cells (MaSC) with contour plots shown for LD (black) and JL (red) tumors. b. Frequency of mammary stem cells in LD and JL tumors. c. Mammosphere-formation efficiency of LD and JL tumor cells. (Source: Hadadi et al., 2019)
Additional data suggest that CRD promotes stemness of primary tumor cells. (By stemness, we mean the ability of a cell to generate differentiated daughter cells and continue its lineage.) The researchers looked at the expression of known markers of mammary cancer-cell stemness and found a significant upregulation of genes associated with the epithelial-mesenchymal transition (EMT), a process where epithelial cells essentially gain the properties to become stem cells. By performing a mammosphere-formation assay—which tests stem cell activity in mammary tissue—the researchers found that mammosphere-formation efficiency (MFE) was much higher in cancer cells from the primary tumors of the jetlag mice (Figure 2). The jetlag mice also showed a decrease in Per2 expression, a gene that increases mammary epithelial and cancer cell stemness. Furthermore, the stemness of the mammary epithelial cells was actually regulated by circadian oscillations of the clock genes. There was a negative correlation between the peaks of PER genes, which peak in the dark, and MFE. Grafted cancer cells purified from the jetlag mice showed an increased tumor-initiating potential in immunocompetent wild-type mice compared to cells from the control mice.
Using flow cytometry, the data show there are reduced numbers of CD45+ immune cells in tumors in the jetlag mice, but there was no major alteration in the proportional distribution of various types of immune cells. However, tumors from jetlag mice had a significantly higher proportion of MCH II-low tumor-associated macrophages (TAMs), which support tumor growth. The jetlag mice also showed high numbers of immunosuppressive CD4+FoxP3+Treg and an elevation of the Treg/CD8 and CD4/CD8 T cell ratios, which are indicators of therapy responsiveness and breast cancer survival. Chronic CRD weakens anti-tumor responses and promotes an immunosuppressive pro-tumor microenvironment. This environment may promote the dissemination of mammary cancer cells and the formation of lung metastasis.
Next, a polymerase chain reaction (PCR) showed that the most downregulated cytokines/chemokines in the primary tumors from jetlag mice are known to favor an anti-tumor immune response. Consistent with the above flow cytometry data, the most upregulated ones are connected to immunosuppression and tumor progression. They injected an inhibitor for CXCR2—a receptor for CXCL5, which promotes cell metastasis through tumor angiogenesis—into another group of jetlag mice and found a decrease in lung metastasis and the amount of PyMT-positive DCCs in the bone marrow. The CD4/CD8 ratio was much lower. This suggests that using a CXCR2 inhibitor could help limit the effect of jetlag on cancer-cell dissemination and metastasis, but this is merely a conceptual finding, and further studies will definitely be required to analyze its ability as a therapy.
It’d be interesting to do more research into the metabolic implications of CRD. While the researchers found a significant increase in plasma lipid levels in the jetlag mice, they only found minimal differences in weight and insulin levels, which the researchers associate with the timeframe of their study and continuous feeding of the jetlag mice. What are the associations between CRD conditions and weight gain, type 2 diabetes and other conditions associated with inconsistent insulin levels? This would require a longer-term study and a method of feeding the jetlag mice that doesn’t minimize physiological differences between awake and rest cycles.

Figure 3. CRD increases the proportion of cancer stem cells (dark blue) and alters the tumour microenvironment by recruiting myeloid-derived suppressor cells (yellow), which creates a suppressive tumour immune microenvironment (TIME), which could relate to the enhanced CXCL5-CXCR2 axis in the TIME. These effects result in increased dissemination and metastasis in bone marrow and lungs. Inhibition of the CXCR2 axis is able to lighten the effect of CRD and promote anti-tumour activity. (Source: Hadadi et al., 2019)
If there’s one big finding to take away from this study, it’s that CRD leads to enhanced cancer-cell dissemination and metastasis. When clock genes undergo altered expression, there are increased risks of cancer severity and metastasis, particularly as the researchers looked at Per2 and Cry2 clock genes. CRD also enhances the tumor-initiating potential of local cells and creates an immunosuppressive local environment. A graphic shows it better than I can (Figure 3).
Annnnd that’s it for this post. Please make sure to leave a comment below and let us know what you thought of the paper and this write up. Until next time, stay curious!